Ty thể – Wikipedia tiếng Việt

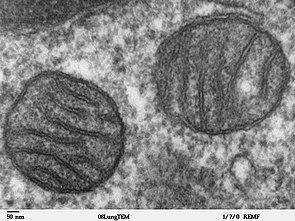 Ảnh chụp hiển vi điện tử của hai ty thể trong tế bào mô phổi động vật hoang dã có vú cho thấy chất nền và những lớp màng bảo phủ bào quan .
Ảnh chụp hiển vi điện tử của hai ty thể trong tế bào mô phổi động vật hoang dã có vú cho thấy chất nền và những lớp màng bảo phủ bào quan .
Ty thể (tiếng Anh: mitochondrion, số nhiều: mitochondria) là một bào quan với màng kép và hiện diện ở tất cả sinh vật nhân thực. Tuy vậy, vẫn có một số tế bào ở các sinh vật đa bào thiếu đi bào quan này (chẳng hạn như tế bào hồng cầu). Một vài sinh vật đơn bào (như Microsporidia, Parabasalia, Diplomonadida) cũng tiêu giảm hoặc biến đổi ty thể của chúng thành những cấu trúc khác.[1] Đến nay, duy chỉ có sinh vật nhân thực chi Monocercomonoides là được biết đã hoàn toàn mất đi ty thể.[2] Trong tiếng Anh, từ mitochondrion bắt nguồn từ tiếng Hy Lạp μίτος, mitos, nghĩa là “sợi” và χονδρίον, chondrion, nghĩa là “hạt”.[3] Ty thể giúp tạo ra phần lớn loại phân tử cao năng là adenosine triphosphate (ATP), một nguồn năng lượng hóa học cung cấp cho hầu hết các hoạt động của tế bào.[4] Chính vì vậy, ty thể còn được gọi là “nhà máy năng lượng của tế bào”.[5]
Đường kính ty thể thường thì giao động từ 0,75 đến 3 μm, [ 6 ] nhưng giữa những ty thể lại độc lạ đáng kể về kích cỡ và cấu trúc. Trừ khi được nhuộm với thuốc nhuộm đặc hiệu, thông thường tất cả chúng ta không hề quan sát được bào quan này. Bên cạnh công dụng cung ứng nguồn năng lượng cho tế bào, ty thể còn tham gia vào những vai trò quan trọng khác, như truyền nhận tín hiệu, biệt hóa tế bào và chết rụng tế bào, cũng như duy trì việc trấn áp chu kỳ luân hồi tế bào và sinh trưởng tế bào. [ 7 ] Những tính năng này được phối hợp một phần bởi quy trình tăng sinh ty thể. [ 8 ] [ 9 ] Ngoài ra, ty thể còn tương quan đến 1 số ít bệnh lý ở người, gồm có rối loạn ty thể, [ 10 ] rối loạn công dụng tim mạch, [ 11 ] suy tim [ 12 ] và tự kỷ. [ 13 ]
Số lượng ty thể trong mỗi tế bào có thể biến động mạnh mẽ tùy thuộc vào từng cơ thể sống, loại mô và loại tế bào. Thí dụ, trong khi tế bào hồng cầu không có ty thể nào, thì tế bào gan lại có thể có hơn 2000 ty thể.[14][15] Ty thể được cấu trúc thành các ngăn hay các phần riêng biệt nhằm đảm nhận những chức năng chuyên hóa khác nhau. Các cấu thành hay được nhắc đến của ty thể bao gồm màng ngoài, xoang gian màng, màng trong, mào và chất nền.
Bạn đang đọc: Ty thể – Wikipedia tiếng Việt
Mặc dù phần nhiều DNA tế bào nằm trong nhân, bào quan ty thể vẫn chiếm hữu một hệ gen độc lập – gần như tựa như hệ gen vi trùng. [ 16 ] Giống như số lượng, protein ty thể cũng dịch chuyển giữa những loại mô và những loài sinh học khác nhau. Ở người, 615 dạng protein khác nhau đã được xác lập từ ty thể của tim, [ 17 ] còn ở chuột, số lượng này là 940 protein. [ 18 ] Hệ protein ty thể được cho rằng có năng lực điều hòa cân đối động. [ 19 ]
Những quan sát đầu tiên về các cấu trúc nội bào có khả năng cao là ty thể đã được công bố vào khoảng thập niên 1840.[20] Năm 1890, Richard Altmann đã chứng minh đây chính là những bào quan tế bào và gọi chúng với cái tên “bioblast” (thể sinh bào).[20][21] Thuật ngữ “mitochondria” (ty thể) được Carl Benda đặt ra 8 năm sau đó.[20][22] Tiếp đến, Leonor Michaelis khám phá ra thuốc nhuộm Janus Green B có thể được sử dụng để nhuộm tươi ty thể vào năm 1900. Năm 1904, Friedrich Meves đã ghi lại những quan sát đầu tiên về ty thể ở thực vật; đối tượng nghiên cứu của ông là tế bào cây hoa súng trắng, Nymphaea alba.[20][23] Năm 1908, ông cùng với Claudius Regaud đề xuất ty thể có thể có chứa protein và lipid. Năm 1912, Benjamin F. Kingsbury lần đầu tiên cho rằng bào quan này có liên quan đến quá trình hô hấp tế bào, nhưng hầu như chỉ dựa trên những quan sát về hình thái học.[20] Một năm sau đó, Otto Heinrich Warburg đã mắc nối những tiểu hạt được tách chiết từ gan chuột với quá trình hô hấp, ông gọi những tiểu hạt đó là “grana”. Một nhà khoa học khác cũng nghiên cứu về vấn đề này là Heinrich Otto Wieland, mặc dù công nhận cơ chế tiểu hạt, ông lại bất đồng với Warburg về bản chất hóa học của quá trình hô hấp. Cuộc tranh cãi vẫn tiếp diễn sau đó, cho đến tận năm 1925 khi David Keilin khám phá ra cytochrome (hay còn gọi là sắc tố tế bào), từ đó hoàn thiện cơ chế chuỗi chuyền điện tử trong hô hấp tế bào.[20]
Năm 1939, những thí nghiệm trên những tế bào cơ tinh chiết đã chứng tỏ rằng : trong quy trình hô hấp tế bào, cứ một nguyên tử oxy đi vào quy trình thì hoàn toàn có thể tạo nên hai phân tử adenosine triphosphate ( ATP ). Đến năm 1941, nhà hóa sinh Fritz Albert Lipmann đã tăng trưởng sáng tạo độc đáo cho rằng những link phosphate của ATP chính là một dạng nguồn năng lượng tham gia vào những quy trình chuyển hóa tế bào. Trong những năm tiếp theo, những chính sách đằng sau quy trình hô hấp tế bào từ từ được triển khai xong, mặc dầu mối liên hệ của quy trình này với bào quan ty thể vẫn chưa được sáng tỏ trọn vẹn. [ 20 ] Kỹ thuật phân đoạn tế bào do Albert Claude trình làng đã được cho phép tách riêng ty thể khỏi những phần còn lại của tế bào và như vậy, ta hoàn toàn có thể nghiên cứu và phân tích hóa sinh chỉ trên riêng đối tượng người tiêu dùng này. Năm 1946, Claude công bố : enzyme cytochrome oxidase cùng những enzyme khác có tham gia vào chuỗi chuyền điện tử trong hô hấp tế bào đã được ông tách chiết từ ty thể. Năm 1948, Eugene Kennedy và Albert Lehninger phát hiện ra ty thể cũng là nơi xảy ra quy trình phosphoryl hóa oxy hóa ở sinh vật nhân thực. Theo thời hạn, công nghệ tiên tiến phân đoạn đã được nâng cấp cải tiến can đảm và mạnh mẽ, nâng cao chất lượng những ty thể phân lập được, cũng như những yếu tố khác tham gia vào quy trình hô hấp tế bào xảy ra tại bào quan này. [ 20 ]Năm 1952, những ảnh chụp vi ký điện tử độ phân giải cao lần tiên phong Open. Phương pháp này dần sửa chữa thay thế kỹ thuật nhuộm bằng thuốc Janus Green cổ xưa và trở thành cách ưa thích để trực quan hóa hình ảnh ty thể. [ 20 ] Phương pháp này giúp những nhà khoa học nghiên cứu và phân tích cấu trúc ty thể một cách chi tiết cụ thể hơn, ví dụ điển hình như xác lập được ty thể có lớp màng bao ngoài. Những tấm ảnh chụp được cũng cho thấy ty thể còn có thêm một lớp màng thứ hai bên trong, gấp nếp thành những mào lấn sâu vào khoang chất nền TT ; đồng thời cũng cho biết sự sai khác đa dạng chủng loại về size và hình dạng ty thể giữa những tế bào với nhau .Biệt danh nổi tiếng của ty thể : ” nhà máy sản xuất nguồn năng lượng của tế bào ” là do Philip Siekevitz đặt cho ty thể năm 1957. [ 24 ]Năm 1967, người ta phát hiện ty thể có chứa ribosome. [ 25 ] Một năm sau đó, 1968, giới khoa học liên tục tăng trưởng những chiêu thức lập map gen ty thể. Cuối cùng, map di truyền và vật lý của hệ gen lập từ DNA ty thể nấm men đã hoàn thành xong năm 1976. [ 20 ]
Nguồn gốc và con đường tiến hóa[sửa|sửa mã nguồn]
Hiện nay có hai giả thuyết về nguồn gốc ty thể : nội cộng sinh và tự sinh. Những yêu cầu từ giả thuyết nội cộng sinh cho rằng ty thể từng là tế bào nhân sơ nguyên thủy, có năng lực triển khai những chính sách oxy hóa mà tế bào nhân thực không hề thực thi được ; và sau đó chúng trở thành những tế bào nội cộng sinh ( endosymbiont ) sinh sống trong những tổ chức triển khai nhân thực. [ 26 ] Còn về giả thuyết tự sinh, người ta cho rằng ty thể sinh ra từ việc chẻ tách một phần DNA của nhân tế bào nhân thực tại thời gian tiến hóa phân hướng khỏi sinh vật nhân sơ ; phần DNA này hoàn toàn có thể được bọc bởi những lớp màng không bị protein pha tạp. Nhưng ngày càng có nhiều dẫn chứng cho thấy ty thể mang nhiều đặc tính chung với vi trùng, do đó dẫn đến việc gật đầu thoáng rộng giả thuyết nội cộng sinh. [ 26 ] [ 27 ]
Mỗi ty thể chứa trong mình vật chất di truyền DNA, gọi là nhiễm sắc thể dạng vòng, được nhân thành một số bản sao từ một phân tử độc nhất. Nhiễm sắc thể ty thể mang các gen quy định protein oxy hóa khử, ví dụ như các enzyme tham gia chuỗi chuyền điện tử hô hấp. Giả thuyết CoRR cho rằng sự đồng định vị này (tức việc những phân tử di truyền định vị trong cùng không gian với các sản phẩm của chính nó) là định chế bắt buộc nhằm điều hòa hoạt động oxy hóa khử trong bào quan. Hệ gen ty thể mã hóa cho một số RNA cấu thành nên ribosome, và 22 loại tRNA cần thiết cho quá trình phiên mã RNA thông tin thành protein. Cấu trúc dạng vòng này cũng có mặt ở sinh vật nhân sơ. Có lẽ vì vậy mà nguyên ty thể (proto-mitochondrion) có thể có mối liên hệ khăng khít với chi Rickettsia.[28][29] Tuy nhiên, thực chất thì tổ tiên ty thể lại có quan hệ mật thiết với lớp Alphaproteobacteria, và người ta đặt ra nghi vấn liệu rằng ty thể được hình thành song hành hay tiếp sau sự xuất hiện của nhân tế bào, vấn đề này vẫn còn trong vòng tranh cãi.[30]
Một điều tra và nghiên cứu mới gần đây [ 31 ] từ những nhà nghiên cứu thuộc Viện Đại học Hawaii tại Manoa và Đại học Tiểu bang Oregon chỉ ra nhánh SAR11 ( bộ Pelagibacterales thuộc lớp Alphaproteobacteria ) của vi trùng năng lực có chung tổ tiên tương đối gần đây với những ty biểu lộ tồn ở hầu hết tế bào nhân thực .
DNA ty thể mã hóa tổng hợp nên những ribosome ty thể, tương tự ribosome vi khuẩn về kích thước và cấu trúc.[33] Nhóm ribosome này giống nhất với ribosome 70S của vi khuẩn và không thuộc nhóm ribosome 80S trôi nổi ngoài tế bào chất do DNA nhân mã hóa.
Mối quan hệ nội cộng sinh giữa ty thể với tế bào chủ chứa chúng theo Lynn Margulis là vô cùng thông dụng. [ 34 ] Thuyết nội cộng sinh đề xuất kiến nghị rằng ty thể là hậu duệ vi trùng, và bằng cách nào đó nó đã sống sót qua quy trình nhập bào đến một tế bào khác, rồi từ từ hòa hợp vào nguyên sinh chất tế bào đó. Khả năng những dạng vi trùng này đã chu toàn tiến trình hô hấp hiếu khí trong tế bào chủ chứa chúng, lúc mà tế bào chủ chỉ sống dựa vào quy trình đường phân và lên men có hiệu suất nguồn năng lượng kém, từ đó tạo ra một lợi thế tiến hóa đáng kể. Quan hệ nội cộng sinh này hoàn toàn có thể đã tăng trưởng từ 1,7 [ 35 ] đến 2 [ 36 ] tỷ năm về trước .Một vài nhóm sinh vật nhân thực đơn bào chỉ còn lại dạng ty thể tiêu giảm hay sót lại những cấu trúc dẫn xuất, ví dụ : ngành Microsporidia, ngành Metamonad và lớp Archamoebae. [ 37 ] Các nhóm này thuộc về những sinh vật nhân thực nguyên thủy nhất nằm trên cây phát sinh chủng loại kiến thiết xây dựng từ thông tin rRNA, và người ta từng cho rằng chúng xuất hiện trước cả lúc ty thể khởi đầu Open. Tuy nhiên, thời nay những nhà khoa học cho đây chỉ là hệ quả từ lỗi mê hoặc nhánh dài ( long branch attraction ) trong phát sinh học. Thực chất chúng lại là những nhóm sinh vật dẫn xuất và giữ lại những gen hay bào quan nguồn gốc từ ty thể ( ví dụ : mitosome và hydrogenosome ). [ 1 ]
Chi Monocercomonoides dường như đã hoàn toàn mất đi ty thể và hiện tại chỉ còn lại một ít chức năng ty thể có khả năng do những protein tế bào chất đảm nhận.[38]
 Siêu cấu trúc ty thể (hình tương tác) Ty thể có một lớp màng kép; trong đó màng trong chứa đựng những bộ máy [39] và xoang gian màng của chúng cũng tương đối mỏng.Ty thể có một lớp màng kép ; trong đó màng trong tiềm ẩn những cỗ máy hóa thẩm và cấu trúc gấp nếp thành những rãnh sâu giúp ngày càng tăng diện tích quy hoạnh mặt phẳng. Bình thường ty thể được miêu tả như ” một chiếc xúc xích cam lỏng chỏng bọng nước bên trong ” ( tựa như hình trên ), thực tiễn thì bào quan này lại sống sót dưới nhiều hình dạng khác nhauvà xoang gian màng của chúng cũng tương đối mỏng mảnh .Mỗi ty thể đều có màng ngoài và màng trong cấu trúc từ lớp phospholipid kép cùng protein. [ 14 ] Mỗi lớp màng lại có những đặc tính khác nhau. Nguyên do phương cách tổ chức triển khai cấu trúc theo hệ màng kép, ty thể được phân thành 5 bộ phận tách biệt. Bao gồm :
Siêu cấu trúc ty thể (hình tương tác) Ty thể có một lớp màng kép; trong đó màng trong chứa đựng những bộ máy [39] và xoang gian màng của chúng cũng tương đối mỏng.Ty thể có một lớp màng kép ; trong đó màng trong tiềm ẩn những cỗ máy hóa thẩm và cấu trúc gấp nếp thành những rãnh sâu giúp ngày càng tăng diện tích quy hoạnh mặt phẳng. Bình thường ty thể được miêu tả như ” một chiếc xúc xích cam lỏng chỏng bọng nước bên trong ” ( tựa như hình trên ), thực tiễn thì bào quan này lại sống sót dưới nhiều hình dạng khác nhauvà xoang gian màng của chúng cũng tương đối mỏng mảnh .Mỗi ty thể đều có màng ngoài và màng trong cấu trúc từ lớp phospholipid kép cùng protein. [ 14 ] Mỗi lớp màng lại có những đặc tính khác nhau. Nguyên do phương cách tổ chức triển khai cấu trúc theo hệ màng kép, ty thể được phân thành 5 bộ phận tách biệt. Bao gồm :
- màng ty thể ngoài
- xoang gian màng (khoảng không gian giữa màng ngoài và màng trong)
- màng ty thể trong
- xoang mào (tạo thành từ sự gấp nếp màng trong), và
- chất nền (không gian bên trong màng trong)
Ty thể nào bị tước đi màng ngoài thì gọi là mitoplast .
Màng ty thể ngoài là lớp màng bao bọc bào quan, độ dày dao động từ 60 đến 75 ångström (Å). Tỷ lệ protein trên phospholipid của màng tương tự màng sinh chất tế bào nhân thực (khoảng 1:1 về mặt khối lượng). Màng ngoài chứa một lượng lớn protein xuyên màng gọi là porin. Những porin tạo nên các kênh cho phép những phân tử nặng khoảng 5000 dalton hoặc nhẹ hơn khuếch tán tự do theo một hướng xác định (từ ngoài vào trong màng hay ngược lại).[14] Protein kích thước lớn có thể xâm nhập ty thể nếu như trình tự tín hiệu tại đầu N liên kết với một phân tử protein lớn giàu tiểu đơn vị gọi là translocase thuộc màng ngoài, để từ đó kích hoạt sự vận chuyển chủ động đưa phân tử xuyên qua màng.[40] Tiền protein ty thể được nhập bào thông qua những phức hệ vận chuyển đặc hiệu. Màng ngoài đồng thời cũng chứa các enzyme tham gia vào nhiều hoạt động đa dạng, như kéo dài phân tử axit béo, oxy hóa hormone adrenaline và phân hủy amino acid tryptophan. Bao gồm những enzyme sau: monoamine oxidase, rotenone-insensitive NADH-cytochrome c-reductase, kynurenine hydroxylase và axit béo Co-A ligase. Sự đánh thủng màng ty thể ngoài dẫn đến hậu quả rò rỉ những protein từ xoang gian màng thấm vào bào tương, gây ra cái chết chắc chắn cho tế bào.[41] Màng ngoài ty thể cũng có khả năng thông kết với những lớp màng của lưới nội chất (endoplasmic reticulum, ER), tạo thành một cấu trúc gọi là MAM (mitochondria-associated ER-membrane, tạm dịch: màng ER liên hợp ty thể). Cấu trúc này đóng vai trò quan trọng trong hoạt động truyền dẫn tín hiệu calci liên ER-ty thể và liên quan đến việc chuyển vận lipid giữa ER và ty thể.[42] Phía ngoài màng ngoài trôi nổi những hạt nhỏ đường kính 60 Å, được đặt tên là tiểu đơn vị Parson (sub-unit of Parson).
Xoang gian màng[sửa|sửa mã nguồn]
Xoang gian màng là khoảng trống choán giữa màng ngoài và màng trong. Đồng thời cũng là khoảng trống bọc quanh ty thể ( perimitochondrial space ). Vì màng ngoài được cho phép những phân tử nhỏ thuận tiện khuếch tán tự do, nên nồng độ của những phân tử này, như ion và đường, ở xoang gian màng tựa như tại bào tương. [ 14 ] Trong khi đó, những protein lớn lại cần phải có trình tự tín hiệu đặc hiệu mới được luân chuyển xuyên thấu màng ngoài, vì thế hàm lượng protein tại xoang gian màng có sự độc lạ so với ngoài bào tương. Ví dụ một loại protein tập trung chuyên sâu trong xoang theo cách này có tên là cytochrome c. [ 41 ]
Màng ty thể trong chứa nhiều loại protein, chia thành 5 nhóm sau theo chức năng từng loại:[14]
Màng trong mang hơn 151 loại polypeptide khác nhau, và có tỷ suất rất cao protein trên phospholipid ( hơn 3 : 1 về mặt khối lượng, nghĩa là cứ khoảng chừng 1 protein thì tương tự với 15 phân tử phospholipid ). Màng trong cũng là bộ phận chiếm giữ xê dịch 1/5 tổng lượng protein ở ty thể. [ 14 ] Ngoài ra, màng còn là nơi tích góp đậm đặc một dạng phospholipid hiếm gặp, cardiolipin. Phân tử phospholipid này khởi đầu được phát hiện trong tim bò năm 1942, và sự xuất hiện của cardiolipin thường là một đặc trưng của màng sinh chất ty thể và vi trùng. [ 43 ] Cardiolipin cấu trúc từ bốn axit béo thay vì hai như thường thì, và hoàn toàn có thể điều này đã khiến cho màng trong trở nên không thấm. [ 14 ] Không như màng ngoài, màng trong lại không có những porin và mang đặc tính không thấm cao so với mọi phân tử muốn chuyển vận xuyên màng. Hầu hết toàn bộ ion và phân tử đều phải nhờ một hệ luân chuyển qua màng đặc hiệu mới hoàn toàn có thể đi vào hay ra khỏi chất nền. Riêng những protein phải được chuyên chở trải qua phức hệ translocase màng trong ( translocase of the inner membrane, TIM ) hay qua Oxa1. [ 40 ] Bên cạnh đó, một điện thế màng cũng Open hai bên màng trong do những ảnh hưởng tác động từ những enzyme thuộc chuỗi chuyền điện tử .
 Hình ảnh cắt ngang của mào trong ty thể gan chuột biểu lộ cấu trúc 3D của nó cũng như mối liên hệ với màng trong
Hình ảnh cắt ngang của mào trong ty thể gan chuột biểu lộ cấu trúc 3D của nó cũng như mối liên hệ với màng trong
Màng ty thể trong được gấp nếp lấn sâu vào chất nền tạo thành nhiều mấu lồi dạng răng lược gọi là mào (cristae, số ít: crista), giúp màng trong mở rộng diện tích bề mặt, nâng cao khả năng sản xuất ATP. Đối với ty thể tế bào gan điển hình, diện tích màng trong lớn gấp 5 lần màng ngoài. Tỷ lệ này cũng biến thiên đa dạng, và đối với những ty thể trong các tế bào có nhu cầu năng lượng ATP cao như tế bào cơ thì chúng lại gấp nếp mạnh mẽ tạo nên nhiều mào hơn. Những nếp màng này được khảm bằng hàng loạt thể cầu nhỏ, gọi là hạt F1 hay oxysome. Đây không hẳn là những nếp gấp đơn giản tạo ra khi màng trong lõm vào chất nền, mà chúng còn có khả năng tác động đến tổng thể chức năng hóa thẩm thấu.[44]
Một nghiên cứu và điều tra mô phỏng toán học gần đây yêu cầu rằng những đặc tính quang học của mào trong ty thể dạng sợi hoàn toàn có thể ảnh hưởng tác động đến sự phát sinh và Viral ánh sáng trong mô. [ 45 ]
Chất nền là khoảng trống bao bởi màng trong. Chứa đựng khoảng chừng 2/3 tổng lượng protein ở ty thể. [ 14 ] Chất nền có vai trò quan trọng trong quy trình sản xuất ATP trải qua mạng lưới hệ thống phức tạp ATP synthase đặt tại màng trong. Đây là nơi đặc trưng bởi nồng độ cao của hỗn hợp hàng trăm enzyme, nhiều ribosome ty thể, tRNA và một số ít bản sao từ hệ gen DNA ty thể. Hoạt động chính của những enzyme chất nền là oxy hóa pyruvate và axit béo, cũng như tham gia quy trình axit citric. [ 14 ]
Ty thể sở hữu vật chất di truyền của riêng chúng, và là bộ máy cấu tạo nên hệ RNA và protein riêng ty thể (xem: sinh tổng hợp protein). Người ta cho biết một trình tự DNA ty thể ở người được công bố chứa đến 16.569 cặp base mã hóa trong 37 gen, gồm: 22 gen tRNA, 2 gen rRNA và 13 gen peptide.[46] Ở người, 13 loại peptide ty thể này khảm vào lớp màng trong, cùng với những protein mã hóa bởi những gen trong nhân tế bào chủ.
Màng ER phối hợp ty thể ( MAM )[sửa|sửa mã nguồn]
Màng lưới nội chất liên hợp ty thể hay màng ER liên hợp ty thể (mitochondria-associated ER membrane, MAM) là thành phần cấu trúc ngày càng được công nhận rộng rãi về vai trò quan trọng của nó trong sinh lý học tế bào và cân bằng nội môi. Từng bị coi là một trở ngại kỹ thuật trong công nghệ phân đoạn tế bào, vì những túi ER pha tạp này khi đó sẽ gây tập nhiễm khi thực hiện phân đoạn tế bào, thì nay được tái xác nhận là một cấu trúc màng bao nguồn gốc từ hệ MAM—phần trung gian chuyển tiếp giữa ty thể và lưới nội chất (ER).[47] Người ta từng quan sát mối liên thông vật lý giữa hai bào quan đây trong những ảnh vi ký điện tử và gần đây với những ảnh chụp hiển vi huỳnh quang.[47] Những nghiên cứu này ước lượng màng ngoài ty thể có khả năng chiếm đến 20% trong tổng cấu trúc MAM, trong đó ER cùng ty thể bị phân cách một khoảng từ 10–25 nm và kết gặp với nhau thông qua những phức hệ buộc nối protein.[47][48][49]
Tinh chế MAM từ quy trình phân đoạn hạ tế bào đã cho thấy cấu trúc này có vai trò làm giàu những enzyme tham gia những phản ứng biến chuyển phospholipid, cạnh bên đó còn là những kênh hoạt động giải trí tương giao với tín hiệu Ca2 +. [ 47 ] [ 49 ] Các tín hiệu trên đã chứng tỏ MAM có một vai trò điển hình nổi bật trong việc điều hòa hoạt động giải trí tàng trữ lipid tế bào và dẫn truyền tín hiệu, kèm với những tất suy ý nghĩa cho hiện tượng kỳ lạ phối hợp ty thể của tế bào, như đoạn trình diễn sau đây. MAM không chỉ phân phối cách nhìn thâm thúy đến những nền tảng cơ học cơ sở như những quy trình sinh lý trong tiến trình tự chết của tế bào cũng như hoạt động giải trí truyền dẫn tín hiệu calci, mà nó còn phác ra một góc nhìn tinh xảo hơn về bào quan ty thể. Dù thường bị xem là tĩnh tại, ” nhà máy sản xuất nguồn năng lượng ” cô lập này lại đoạt chiếm hoạt động giải trí chuyển hóa của tế bào qua một sự kiện nội cộng sinh nguyên thủy, do vậy con đường tiến hóa của MAM phải gạch ra những số lượng giới hạn để ty thể được phép tích hợp hòa hợp vào toàn diện và tổng thể quy trình sinh lý tế bào, kèm theo những khớp buộc vật lý và nhập hòa công dụng với mạng lưới hệ thống nội màng .
Vận chuyển phospholipid[sửa|sửa mã nguồn]
MAM có năng lực làm giàu những enzyme tham gia sinh tổng hợp lipid, như phosphatidylserine synthase trên mặt phẳng lưới nội chất và phosphatidylserine decarboxylase trên mặt phẳng ty thể. [ 50 ] [ 51 ] Nguyên do ty thể là bào quan liên tục dung hợp và phân đôi, nên chúng nhu yếu phải có một nguồn cung ứng phospholipid liên tục và chất lượng nhằm mục đích duy trì bộ màng được nguyên vẹn. [ 52 ] [ 53 ] Nhưng ty thể không chỉ là đích nhập phospholipid tại tận cùng tiến trình tổng hợp ; mà đúng hơn, bào quan này còn đóng vai trò trong hệ liên bào quan để giao thương mua bán những chất trung gian và những mẫu sản phẩm từ con đường sinh tổng hợp phospholipid, chuyển hóa ceramide và cholesterol, cũng như đồng nhất glycosphingolipid. [ 51 ] [ 53 ]
Việc MAM điều tiết khả năng giao thương nội bào được chứng minh là để tạo điều kiện thuận lợi cho sự chuyển vận lipid trung gian giữa các bào quan.[50] Tương phản với cơ chế chuyển tiếp lipid căn bản qua hệ túi sinh học, bằng chứng cho thấy khoảng cách vật lý gần giữa màng ER và ty thể tại MAM cho phép lipid dịch chuyển linh hoạt giữa hai lớp màng đối diện nhau này.[53] Dẫu cho cơ chế đây lại hiếm gặp và dường như thu năng lượng, như chất vận chuyển không yêu cầu ATP.[53] Thay vì đó, ở nấm men, sự vận chuyển này lại phụ thuộc vào một cấu trúc buộc kết đa protein gọi là cấu trúc gặp gỡ ER-ty thể (ER-mitochondria encounter structure, hay ERMES), mặc dù vẫn chưa rõ liệu cấu trúc này có trực tiếp làm trung gian vận chuyển lipid hay chỉ có vai trò cần thiết để neo giữ các màng ở vị trí đủ gần nhằm tiêu giảm hàng rào năng lượng cho hoạt động lipid dịch chuyển linh hoạt.[53][54]
MAM đồng thời cũng hoàn toàn có thể là thành phần thuộc lộ trình chế tiết ( secretory pathway ), bên cạnh vai trò trong giao thương mua bán lipid nội bào. Cụ thể, MAM có vẻ như là một trạm trung chuyển giữa lưới nội chất hạt ( rough ER ) và cỗ máy Golgi trên lộ trình, từ đó hướng dẫn cho những lipoprotein tỷ trọng rất thấp ( very-low-density lipoprotein, hay VLDL ) lắp rắp hoặc bài tiết ra ngoài. [ 51 ] [ 55 ] Như vậy, MAM chính là một TT giao thương mua bán và trao đổi chất quan trọng trong quy trình chuyển hóa lipid .
Tín hiệu calci[sửa|sửa mã nguồn]
Vai trò then chốt của ER trong phát xuất tín hiệu calci được công nhận trước khi người ta gật đầu thoáng đãng ty thể cũng có vai trò trong quy trình này, một phần là do đặc tính ái lực thấp của những kênh Ca2 + xác định tại màng ty thể ngoài có vẻ như đối ngược với năng lực phân phối có mục tiêu của bào quan này nhằm mục đích tạo nên những đổi khác trên dòng chảy Ca2 + nội bào. [ 47 ] [ 56 ] Nhưng sự hiện hữu của MAM đã giúp xử lý xích míc rõ ràng đây : sự phối hợp vật lý khoảng cách gần giữa hai bào quan dẫn đến hệ quả những vi miền Ca2 + ( Ca2 + microdomain ) tại những điểm tiếp xúc hoàn toàn có thể thuận tiện truyền tải hiệu suất cao Ca2 + từ ER đến ty thể. [ 47 ] Quá trình truyền dẫn xảy ra khi phân phối lại thứ gọi là ” điểm phình Ca2 + ” ( Ca2 + puff ), tạo từ việc nhóm cụm tự phát và kích hoạt công suất phức hệ IP3R, một kênh Ca2 + xuyên màng ER tiêu chuẩn. [ 47 ] [ 48 ]Số phận của những điểm phình này — đơn cử, liệu rằng chúng vẫn hạn chế chuyển đến những khu vực khác biệt hay tích kết vào những cơn sóng Ca2 + lưu thông khắp tế bào — được định rõ ở một toàn diện và tổng thể lớn hơn theo những hoạt độ động lực của MAM. Dù cho sự tái hấp thu Ca2 + của ER ( đồng thời với sự phát xuất ) điều biến cường độ điểm phình, như vậy cũng gây cách điện ty thể ở một mức độ nhất định từ việc phơi nhiễm nồng độ cao Ca2 +, MAM thường cư xử như một tường lửa, về mặt thực chất là những điểm phình Ca2 + kèm đệm, bằng cách chuyển thành một bể chìm để những ion tự do giải phóng vào bào tương hoàn toàn có thể được phễu lọc. [ 47 ] [ 57 ] [ 58 ] Đường hầm Ca2 + Open trải qua thụ quan Ca2 + ái lực thấp là VDAC1, gần đây được chứng tỏ là hoàn toàn có thể buộc kết về mặt vật lý với những cụm IP3R trên màng ER và được làm giàu tại MAM. [ 47 ] [ 48 ] [ 59 ] Khả năng ty thể hành xử như một bể chứa Ca2 + là một hệ quả từ gradient điện hóa Open xuyên suốt quy trình phosphoryl hóa oxy hóa, tạo nên những đường hầm cation theo tiến trình sụt thế. [ 59 ] Thông thường, dòng calci trung hòa từ bào tương chảy vào chất nền ty thể là nguyên do gây ra hiện tượng kỳ lạ khử cực trong thời điểm tạm thời, rồi được hiệu sửa sau đó bằng cách bơm những proton ra ngoài .Lưu ý là quy trình truyền dẫn Ca2 + không phải một chiều đơn nhất ; đúng chuẩn hơn là con đường lưu thông hai chiều. [ 56 ] Những đặc thù của bơm Ca2 + SERCA và kênh IP3R bộc lộ trên màng ER tạo điều kiện kèm theo xảy ra sự điều hòa ngược điều phối nhờ công dụng của MAM. Cụ thể, việc MAM quét sạch Ca2 + cho phép mẫu hóa không thời hạn ( spatio-temporal patterning ) so với sự phát xuất tín hiệu Ca2 + bởi Ca2 + hoàn toàn có thể tu chỉnh hoạt động giải trí IP3R một cách lưỡng pha. [ 47 ] SERCA cũng chịu ảnh hưởng tác động tựa như từ tác động ảnh hưởng ngược của ty thể : hấp thu Ca2 + khi MAM kích thích sản xuất ATP, như vậy cung ứng nguồn năng lượng được cho phép SERCA nạp tải Ca2 + qua ER từ những dòng Ca2 + tuôn trào liên hồi tại MAM. [ 57 ] [ 59 ] Như thế, MAM không chỉ là bộ đệm thụ động của những điểm phình Ca2 +, mà còn giúp điều tiết những dòng tín hiệu Ca2 + xa hơn nữa trải qua những vòng lặp hồi ngược tác động ảnh hưởng đến hoạt tính động lực của ER .Quá trình điều hòa ER giải phóng Ca2 + tại MAM đặc biệt quan trọng quan trọng bởi hoàn toàn có thể chỉ có một lối thông nhất định hấp thu Ca2 + nhằm mục đích duy trì hoạt động giải trí ty thể, và ở mức độ tế bào là quy trình hóa thẩm thấu. Một lượng vừa đủ tín hiệu Ca2 + nội bào quan là nhu yếu thiết yếu để kích thích trao đổi chất bằng cách hoạt hóa những enzyme dehydrogenase có vai trò chu toàn quy trình axit citric. [ 60 ] Tuy nhiên, một khi lượng tín hiệu Ca2 + trong ty thể vượt ngưỡng nhất định lại dẫn đến kích hoạt phương pháp nội tại của quy trình chết rụng tế bào, một phần nguyên do sự suy biến thế năng màng ty thể thiết yếu cho hoạt động giải trí trao đổi chất. [ 47 ] Những nghiên cứu và điều tra khảo sát vai trò những tác nhân tiền và phản chết rụng ( pro – and anti-apoptotic factor ) đã ủng hộ quy mô này, lấy ví dụ, tác nhân phản chết rụng Bcl-2 được dẫn chứng là hoàn toàn có thể tương tác với IP3R nhằm mục đích hạ lượng Ca2 + chảy vào ER, dẫn đến tiêu giảm dòng chảy tuôn trào tại MAM và ngăn ngừa sự suy biến thế năng màng ty thể do chất kích thích hậu chết rụng ( post-apoptotic stimuli ). [ 47 ] Vì nhu yếu điều hòa khỏe mạnh dòng tín hiệu Ca2 +, có lẽ rằng không quá bất ngờ mấy khi sự rối loạn điều hòa Ca2 + ty thể lại tương quan đến một số ít bệnh lý thoái hóa thần kinh, khi mà hàng loạt gen ức chế khối u bao hàm một vài phần mã hóa cho những loại sản phẩm được làm giàu tại MAM. [ 59 ]
Cơ sở phân tử của chính sách buộc kết[sửa|sửa mã nguồn]
Những văn minh gần đây trong việc nhận diện cấu trúc buộc kết giữa màng ty thể và ER đã gợi ý công dụng giàn giáo của những thành phần phân tử là thứ sinh so với những tính năng không tham gia cấu trúc của chúng. Ở nấm men, ERMES, một phức hệ đa protein cư trú liên màng ER và ty thể, bắt buộc phải có cho hoạt động giải trí luân chuyển lipid tại MAM và cũng là thành phần minh họa cho nguyên tắc trên. Lấy ví dụ, một trong những cấu phần của nó cũng là một cấu thành của hệ phức tạp protein cần có cho sự chèn gắn những protein ống beta xuyên màng ( transmembrane beta-barrel protein ) vào lớp lipid kép. [ 53 ] Tuy vậy, một đồng đẳng của phức hệ ERMES vẫn chưa được xác lập trọn vẹn trong tế bào động vật hoang dã có vú. Những protein trực tiếp khác trong hệ cấu trúc giàn giáo cũng có tựa như những tính năng độc lập với hoạt tính buộc kết có cấu trúc tại MAM ; tỷ dụ, những mitofusin cư trú liên ER và ty thể hình thành nên những dị phức hệ ( heterocomplex ) quản trị một lượng vị trí tiếp túc liên bào quan, mặc dầu người ta chỉ mới xác lập được vai trò của mitofusin trong những sự kiện phân đôi và dung hợp giữa những ty thể riêng không liên quan gì đến nhau. [ 47 ] Protein điều hòa glucose 75 ( Glucose-regulated protein 75, Grp75 ) là một protein tuy nhiên năng khác. Bên cạnh việc tập trung chuyên sâu thành những bể Grp75 trong chất nền, một phần phân tử cũng đảm trách như một chaperone trong việc link vật lý giữa những kênh Ca2 + VDAC và IP3R liên ty thể-ER nhằm mục đích nâng cao hiệu suất cao truyền dẫn Ca2 + tại MAM. [ 47 ] [ 48 ] Ngoài ra còn có một cấu trúc buộc kết thế năng nữa là Sigma-1R, một thụ quan phi opioid ( non-opioid receptor ) có công dụng không thay đổi hóa phức hệ IP3R cư trú ER, có năng lực bảo tồn thông tin liên lạc tại MAM trải qua những cung ứng stress chuyển hóa. [ 61 ] [ 62 ]
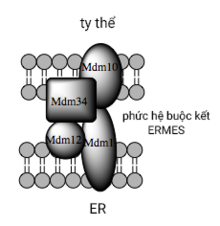 Mô hình phức hệ buộc kết đa protein ở nấm men, ERMES
Mô hình phức hệ buộc kết đa protein ở nấm men, ERMES
MAM là một TT tín hiệu, chuyển hóa và giao thương mua bán quan trọng trong tế bào, đảm nhiệm sự phối hợp sinh lý giữa ER và ty thể. Những khớp buộc giữa hai bào quan này không chỉ đơn thuần là một cấu trúc thông thường mà còn chịu nghĩa vụ và trách nhiệm triển khai nhiều công dụng có ích và quan trọng trong toàn diện và tổng thể tiến trình sinh lý tế bào cũng như cân đối nội môi. Như vậy, MAM đã áp thêm một quan điểm mới lên ty thể phân tán khỏi cách nhìn truyền thống lịch sử về bào quan này như một thể tĩnh tại, một đơn vị chức năng cô lập từng đoạt chiếm năng lực chuyển hóa của tế bào. Thay cho ý niệm cũ đó, thực tiễn chính cấu trúc liên ER-ty thể này đã nhấn mạnh vấn đề tính phối hợp của ty thể, loại sản phẩm từ một sự kiện nội cộng sinh, trong những quy trình tế bào phong phú .
Tổ chức và phân bổ[sửa|sửa mã nguồn]
 Mạng lưới ty thể nổi bật ( xanh lục ) trong hai tế bào người ( tế bào HeLa
Mạng lưới ty thể nổi bật ( xanh lục ) trong hai tế bào người ( tế bào HeLa
Ty thể (cùng những cấu trúc liên quan) có mặt ở tất cả sinh vật nhân thực (trừ một trường hợp duy nhất—chi Monocercomonoides thuộc bộ Oxymonadida).[2][63] Về mặt tổ chức, ty thể hình thành nên một mạng lưới năng động ở phần lớn tế bào nơi chúng liên tục phân đôi và dung hợp. Số lượng và khu vực tập trung bào quan này cũng thay đổi đa dạng tùy vào loại tế bào. Một ty thể đơn thường chỉ tìm thấy trong những sinh vật đơn bào. Tương phản với số lượng lớn ty thể nằm trong các tế bào gan người, tức khoảng 1000–2000 ty thể trên một đơn vị tế bào, chiếm đến 1/5 thể tích tế bào.[14] Mặt khác hàm lượng ty thể trong những tế bào tương tự cũng biến thiên đáng kể tùy vào kích thước và thế năng màng,[64] với những chênh lệch phát sinh từ căn nguyên bao gồm cả việc phân chia nguyên sinh chất không đồng đều khi phân bào, dẫn đến những sai khác ngoại lai tác động đến nồng độ ATP và các quá trình tế bào thuận chiều.[65] Ty thể có thể bị kẹp giữa những sợi cơ (myofibril) của cơ hay bọc quanh roi tinh trùng.[14] Bình thường, chúng cấu thành một mạng lưới tỏa nhánh 3 chiều phức tạp neo cùng hệ khung xương tế bào. Sự liên hợp với bộ khung xương giúp xác định nên hình dạng ty thể, từ đó có khả năng ảnh hưởng đến nhóm chức năng đặc hiệu:[66] những cấu trúc khác nhau của mạng lưới ty thể có lẽ đủ khả năng để tạo nên một quần tụ đa dạng ưa nhược các đặc tính vật lý, hóa học và tín hiệu.[67] Trong tế bào, ty thể luôn được phân phối dọc theo hệ thống vi ống và sự phân phát bào quan này cũng có mối liên hệ với mạng lưới nội chất.[68] Những bằng chứng gần đây đề xuất rằng vimentin, một trong nhiều thành phần của khung xương tế bào, cũng có vai trò quan trọng trong hiện tượng liên hợp trên.[69]
Vai trò điển hình nổi bật nhất của ty thể là sản sinh ” đồng xu tiền nguồn năng lượng ” của tế bào, chính là phân tử cao năng ATP ( tức dạng phosphoryl hóa của ADP ), trải qua quy trình hô hấp tế bào ; và đồng thời điều hòa hoạt động giải trí trao đổi chất tế bào. [ 15 ] Phản ứng TT trong tổng thể và toàn diện tiến trình sản xuất ATP chung quy lại là quy trình axit citric, hay còn gọi quy trình Krebs. Ngoài ra, ty thể còn tiếp đón hàng loạt công dụng khác bên cạnh vai trò sản xuất ATP .
Chuyển hóa nguồn năng lượng[sửa|sửa mã nguồn]
Vai trò chính yếu của ty thể là sản xuất ATP, biểu lộ qua một lượng lớn protein tại màng trong Giao hàng công dụng này. Các protein này sản sinh ATP từ mẫu sản phẩm oxy hóa chính của glucose là pyruvate, tạo ra trong bào tương cùng với những phân tử NADH. [ 15 ] Kiểu hô hấp tế bào xảy ra ở ty thể có tên gọi hô hấp hiếu khí, phụ thuộc vào vào sự hiện hữu của khí oxy. Khi nồng độ oxy thấp hơn ngưỡng tối thiểu, những mẫu sản phẩm đường phân sẽ bị chuyển hóa qua hoạt động giải trí lên men kỵ khí, một quy trình độc lập với bào quan ty thể. [ 15 ] Sự sản sinh ATP từ glucose trong hô hấp hiếu khí có hiệu suất cao gấp 13 lần trong quy trình lên men. [ 70 ] Gần đây người ta phát hiện ty thể thực vật hoàn toàn có thể sản xuất một lượng hạn chế ATP mà không cần oxy bằng cách sử dụng cơ chất nitrit sửa chữa thay thế. [ 71 ] ATP xuyên qua màng trong với sự trợ giúp của một loại protein đặc hiệu, và đi xuyên màng ngoài trải qua những porin. Tương tự, ADP trở lại cũng vận động và di chuyển theo lộ trình như vậy .
Pyruvate và quy trình axit citric[sửa|sửa mã nguồn]
Những phân tử pyruvate sinh ra từ quy trình đường phân được luân chuyển dữ thế chủ động xuyên qua màng ty thể trong và chuyển đến chất nền, nơi chúng hoàn toàn có thể liên tục bị oxy hóa và hóa hợp với coenzyme A hình thành nên CO2, acetyl-CoA và NADH ; [ 15 ] hoặc bị carboxyl hóa ( bằng enzyme pyruvate carboxylase ) tạo ra oxaloaxetat. Chính phản ứng carboxyl này sẽ giúp ngày càng tăng hàm lượng oxaloacetate trong quy trình axit citric, thế cho nên nó thuộc loại phản ứng bổ trợ ( anaplerotic reaction ), tăng cường hiệu suất quy trình để chuyển hóa mạnh hơn acetyl-CoA khi nhu yếu nguồn năng lượng của mô ( như trong cơ ) bật cao bất thần đang lúc hoạt động giải trí. [ 72 ]Trong quy trình axit citric, tổng thể những chất trung gian ( ví dụ : citrate, iso-citrate, alpha-ketoglutarate, succinate, fumarate, malate và oxaloacetate ) được tái sinh trong suốt mỗi vòng quy trình. Khi bổ trợ nhiều hơn bất kể chất nào trong số những chất trung gian trên vào ty thể đều dẫn đến hiện tượng kỳ lạ hàm lượng bổ trợ chất đó vẫn không mất đi trong quy trình, mà còn làm tăng cao nồng độ những chất trung gian khác một khi nó bị chuyển hóa thành những loại sản phẩm tiếp nối đuôi nhau sau đó. Do vậy, việc thêm vào bất kể chất nào trong số chúng đến quy trình sẽ gây ra một hiệu ứng bổ trợ ( anaplerotic effect ), và ngược lại là hiệu ứng rút bỏ ( cataplerotic effect ). Những phản ứng bổ trợ và rút bỏ này sẽ làm tăng hoặc giảm hàm lượng oxaloacetate sẵn có để hóa hợp với acetyl-CoA hình thành nên axit citric trong suốt lộ trình quy trình. Các sự kiện trên tạo nên những biến chuyển tăng cường hoặc hạ thấp cường độ sản xuất ATP trong ty thể, cũng như ảnh hưởng tác động đến nguồn cung sẵn có ATP cho tế bào. [ 72 ]Mặt khác, acetyl-CoA dù cho sinh ra từ quy trình oxy hóa pyruvate hay từ tiến trình beta-oxy hóa axit béo cũng đều là nguyên vật liệu duy nhất nhập vào quy trình axit citric. Với mỗi vòng quy trình, một phân tử acetyl-CoA sẽ hóa hợp vào bất kể phân tử oxaloacetate nào đang hiện hữu tại chất nền ty thể, và không khi nào được tái sinh trở lại. Đó chính là quy trình oxy hóa phần acetate của acetyl-CoA, tạo ra CO2 và nước, kèm theo nguồn năng lượng phát thải dưới dạng ATP. [ 72 ]Trong gan, sự carboxyl hóa pyruvate bào tương thành oxaloacetate nội ty thể là bước khởi đầu của con đường tân tạo glucose ( gluconeogenic pathway ), để rồi sau đó chuyển hóa thành lactate và alanine dạng khử amine hóa thành glucose, [ 15 ] [ 72 ] tiến trình này chịu ảnh hưởng tác động bởi nồng độ cao glucagon và / hoặc adrenalin trong máu. [ 72 ] Lúc này đây, sự thêm vào oxaloacetate đến ty thể không còn gây ra một hiệu ứng bổ trợ rõ ràng nữa, trong khi một chất trung gian khác của quy trình axit citric là malate thì lại lập tức loại đi khỏi ty thể để chuyển về dạng oxaloacetate bào tương, dẫn đến hệ quả ở đầu cuối là trở thành glucose, một quy trình hầu hết trọn vẹn đảo ngược so với đường phân. [ 72 ]
Những enzyme thuộc chu trình axit citric đắm chìm trong chất nền ty thể, trừ ngoại lệ enzyme succinate dehydrogenase lại khảm quanh màng ty thể trong vì nó là một bộ phận của Phức hệ II (Complex II).[73] Chu trình axit citric oxy hóa acetyl-CoA thành cacbon dioxide, và tạo nên những cofactor dạng khử (3 phân tử NADH và 1 phân tử FADH2), đây là nguồn electron dồi dào cung cấp cho chuỗi chuyền điện tử; cùng với một phân tử GTP (dễ dàng chuyển hóa thành ATP).[15]
NADH và FADH2 : chuỗi chuyền điện tử[sửa|sửa mã nguồn]
![]() Sơ đồ chuỗi chuyền điện tử trong xoang gian màng ty thểNăng lượng oxy hóa từ NADH và FADH2 được chuyển đến oxy ( O2 ) trải qua một vài quy trình tiến độ thuộc chuỗi chuyền điện tử. Các phân tử giàu nguồn năng lượng này sản sinh tại chất nền từ quy trình axit citric và một phần đến từ quy trình đường phân tại tế bào chất. Đương lượng khử từ tế bào chất có năng lực được nhập cảng trải qua mạng lưới hệ thống con thoi malate-aspartate thuộc nhóm protein đối chuyển, hay chuyển vào chuỗi chuyền điện tử bằng cách sử dụng con thoi glycerol phosphate. [ 15 ] Những phức hệ protein tại màng trong ( NADH dehydrogenase ( ubiquinone ), cytochrome c reductase và cytochrome c oxidase ) đảm nhiệm năng lực truyền dẫn và ngày càng tăng giải phóng nguồn năng lượng bằng việc bơm proton ( H + ) xuất ra xoang gian màng. Tổng thể hiệu suất quy trình là hiệu suất cao, nhưng một lượng nhỏ electron hoàn toàn có thể sớm khử oxy, tạo nên những chủng chất hoạt động giải trí có oxy ( reactive oxygen species ) như superoxit. [ 15 ] Hiện tượng này hoàn toàn có thể gây ra căng thẳng mệt mỏi oxy hóa trong ty thể và góp thêm phần suy giảm tính năng ty thể cũng như dẫn đến lão hóa. [ 74 ]Khi tăng nồng độ proton trong xoang gian màng, một gradient điện hóa mạnh được thiết lập hai bên màng trong. Các proton hoàn toàn có thể trở về chất nền trải qua phức hệ ATP synthase, và chính nguồn năng lượng thế năng này sẽ giúp tổng hợp ATP từ ADP và phosphate vô cơ ( Pi ). [ 15 ] Quá trình này được gọi là hóa thẩm thấu ( chemiosmosis ), do nhà khoa học Peter Mitchell [ 75 ] [ 76 ] lần tiên phong phát hiện, ông đã đoạt Giải Nobel hóa học 1978 cho chính tò mò này. Sau này, một phần Giải Nobel hóa học 1997 trao cho Paul D. Boyer và John E. Walker qua những góp sức của về việc giải thuật chính sách hoạt động giải trí ATP synthase. [ 77 ]
Sơ đồ chuỗi chuyền điện tử trong xoang gian màng ty thểNăng lượng oxy hóa từ NADH và FADH2 được chuyển đến oxy ( O2 ) trải qua một vài quy trình tiến độ thuộc chuỗi chuyền điện tử. Các phân tử giàu nguồn năng lượng này sản sinh tại chất nền từ quy trình axit citric và một phần đến từ quy trình đường phân tại tế bào chất. Đương lượng khử từ tế bào chất có năng lực được nhập cảng trải qua mạng lưới hệ thống con thoi malate-aspartate thuộc nhóm protein đối chuyển, hay chuyển vào chuỗi chuyền điện tử bằng cách sử dụng con thoi glycerol phosphate. [ 15 ] Những phức hệ protein tại màng trong ( NADH dehydrogenase ( ubiquinone ), cytochrome c reductase và cytochrome c oxidase ) đảm nhiệm năng lực truyền dẫn và ngày càng tăng giải phóng nguồn năng lượng bằng việc bơm proton ( H + ) xuất ra xoang gian màng. Tổng thể hiệu suất quy trình là hiệu suất cao, nhưng một lượng nhỏ electron hoàn toàn có thể sớm khử oxy, tạo nên những chủng chất hoạt động giải trí có oxy ( reactive oxygen species ) như superoxit. [ 15 ] Hiện tượng này hoàn toàn có thể gây ra căng thẳng mệt mỏi oxy hóa trong ty thể và góp thêm phần suy giảm tính năng ty thể cũng như dẫn đến lão hóa. [ 74 ]Khi tăng nồng độ proton trong xoang gian màng, một gradient điện hóa mạnh được thiết lập hai bên màng trong. Các proton hoàn toàn có thể trở về chất nền trải qua phức hệ ATP synthase, và chính nguồn năng lượng thế năng này sẽ giúp tổng hợp ATP từ ADP và phosphate vô cơ ( Pi ). [ 15 ] Quá trình này được gọi là hóa thẩm thấu ( chemiosmosis ), do nhà khoa học Peter Mitchell [ 75 ] [ 76 ] lần tiên phong phát hiện, ông đã đoạt Giải Nobel hóa học 1978 cho chính tò mò này. Sau này, một phần Giải Nobel hóa học 1997 trao cho Paul D. Boyer và John E. Walker qua những góp sức của về việc giải thuật chính sách hoạt động giải trí ATP synthase. [ 77 ]
Dưới những điều kiện nhất định, các proton có thể nhập lại chất nền ty thể mà không tạo ra ATP. Quá trình này được gọi là rò rỉ proton (proton leak) hay tách cặp ty thể (mitochondrial uncoupling) và nguyên nhân là do hiện tượng khuếch tán tăng cường proton chảy vào chất nền. Từ đó dẫn đến hệ quả rơi tuột năng lượng thế năng của gradient điện hóa proton chuyển thoát thành nhiệt năng.[15] Quá trình trên do một kênh proton tên thermogenin (hay UCP1) đảm nhận vai trò là chất trung gian.[78] Thermogenin là protein nặng 33 kDa, phát hiện lần đầu năm 1973.[79] Kênh này chủ yếu tập trung ở mô mỡ nâu, gọi tắt mỡ nâu, chịu trách nhiệm cho cơ chế sinh nhiệt không run (non-shivering thermogenesis). Mô mỡ nâu có ở động vật có vú và đạt khối lượng cao nhất tại những năm tháng đầu đời và trong những động vật ngủ đông. Ở người, mô mỡ nâu hiện diện khi cơ thể mới sinh và tiêu dần lúc lớn lên.[78]
Lưu trữ ion calci[sửa|sửa mã nguồn]
Nồng độ calci tự do trong tế bào có thể điều biến hàng loạt phản ứng và là thuộc tính quan trọng của cơ chế truyền dẫn tín hiệu nội bào. Ty thể có khả năng lưu trữ calci nhanh chóng, đảm bảo cân bằng nội môi hàm lượng calci trong tế bào.[80][81] Trên thực tế, khả năng chiếm lấy nhanh nhạy calci dự trữ cho lần giải phóng lát sau khiến chúng trở thành “bộ đệm bào tương” rất tốt cho calci.[82][83][84] Lưới nội chất (endoplasmic reticulum, ER) là khu vực lưu trữ calci quan trọng nhất,[56] và cùng với ty thể tạo nên mối tương tác sâu rộng về hoạt động trao đổi calci.[85] Các ion calci phiêu bạt được di lý đến chất nền với sự giúp đỡ của protein đơn chuyển calci ty thể (mitochondrial calcium uniporter) khảm trên màng trong.[86] Cơ chế này chủ yếu do điện thế màng ty thể điều khiển.[81] Sự phóng thích calci trở về không gian nội bào có thể thực hiện thông qua protein trao đổi natri-calci hay qua con đường giải phóng calci cảm ứng calci (calcium-induced calcium release).[86] Sự kiện này có thể khơi mào phát xuất những ghim bắn hay cơn sóng calci kèm theo những thay đổi lớn trên điện thế màng. Từ đó có thể tiếp tục kích hoạt một loạt các protein thuộc hệ thống chất truyền tin cấp hai để phối hợp hoạt động những quá trình song hành với nhau, như giải phóng chất dẫn truyền xung thần kinh trong tế bào thần kinh và giải phóng các hormone trong tế bào nội tiết.[87]
Gần đây người ta cho rằng những luồng Ca2 + chảy đến chất nền ty thể chính là một chính sách điều hòa hoạt động giải trí sinh nguồn năng lượng hô hấp bằng việc cho phép thế điện hóa hiện hữu hai bên màng để ” bật nhảy ” bất ngờ đột ngột từ trạng thái chênh lệch thế năng lợi thế ( ΔΨ-dominated ) sang pH lợi thế ( pH-dominated ), tạo điều kiện kèm theo thuyên giảm stress oxy hóa. [ 88 ] Ở neuron, sự ngày càng tăng nồng độ calci trong bào tương và ty thể bảo vệ không thay đổi hóa hoạt động giải trí thần kinh sát cánh với công dụng chuyển hóa nguồn năng lượng của bào quan này. Hàm lượng calci trong chất nền ty thể hoàn toàn có thể đạt đến hàng chục micromol, đủ ngưỡng thiết yếu để kích hoạt isocitrate dehydrogenase, một trong những enzyme then chốt của quy trình Krebs. [ 89 ]
Những tính năng khác[sửa|sửa mã nguồn]
Ty thể còn đóng vai trò TT trong nhiều tiến trình trao đổi chất khác, ví dụ :
Một vài công dụng của ty thể chỉ hiện hữu ở 1 số ít loại tế bào đặc biệt quan trọng. Thí dụ, ty thể tế bào gan chứa những enzyme được cho phép triển khai giải độc amonia, là loại sản phẩm thải từ con đường chuyển hóa protein. Mỗi một đột biến trong điều hòa biểu lộ gen pháp luật những tính năng trên đều hoàn toàn có thể dẫn đến hậu quả gây ra những bệnh lý ty thể .
Điều hòa tăng sinh tế bào[sửa|sửa mã nguồn]
Mối quan hệ giữa sự tăng sinh tế bào và ty thể đã được điều tra và nghiên cứu trên cơ sở nguồn từ những tế bào ung thư cổ tử cung HeLa. Những tế bào khối u nhu yếu phải chu cấp một lượng lớn ATP để tổng hợp những hoạt chất sinh học như lipid, protein và nucleotide nhằm mục đích tăng sinh tế bào nhanh gọn. [ 98 ] Đa số ATP trong tế bào khối u hình thành từ quy trình phosphoryl hóa oxy hóa ( oxidative phosphorylation, hay OxPhos ). [ 99 ] Như vậy, sự can thiệp đến OxPhos khiến đình chỉ chu kỳ luân hồi tế bào chứng tỏ rằng ty thể góp chân một vai trò trong hoạt động giải trí tăng sinh tế bào. [ 99 ] Ngoài ra, sự sản sinh ATP của ty thể cũng quan trọng trong quy trình phân bào, bên cạnh những công dụng cơ bản khác trong tế bào như lao lý thể tích tế bào, nồng độ dịch bào và kiến trúc tế bào. [ 100 ] [ 101 ] [ 102 ] Hàm lượng ATP giao động khác nhau theo từng pha trong chu kỳ luân hồi tế bào cho thấy mối liên hệ mật thiết giữa dung tích ATP và sức khỏe thể chất tế bào để mở ra một chu kỳ luân hồi tế bào mới. [ 103 ] Vai trò của ATP trong hệ công dụng tế bào cơ bản khiến cho chu kỳ luân hồi tế bào trở nên nhạy cảm với số lượng ATP khả dụng tạo từ ty thể. [ 103 ] Những nồng độ ATP độc lạ sống sót giữa những pha khác nhau của chu kỳ luân hồi tế bào tương hỗ cho giả thuyết định rằng ty thể đóng một vai trò quan trọng trong hoạt động giải trí điều hòa chu kỳ luân hồi tế bào. [ 103 ] Mặc dù những chính sách đặc hiệu trong tương tác giữa ty thể và sự điều hòa chu kỳ luân hồi tế bào vẫn chưa được hiểu rõ toàn vẹn, nhưng những điều tra và nghiên cứu chỉ ra rằng những điểm trấn áp nguồn năng lượng thấp của chu kỳ luân hồi giúp giám sát cường độ nguồn năng lượng trước khi tế bào chạm đến một vòng phân bào mới. [ 104 ]
 Hệ gen ty thể người tiềm ẩn 16.569 cặp base mã hóa trong 37 gen, chia ra 28 gen trên mạch H ( H-strand ) và 9 gen trên mạch L ( L-strand ) .
Hệ gen ty thể người tiềm ẩn 16.569 cặp base mã hóa trong 37 gen, chia ra 28 gen trên mạch H ( H-strand ) và 9 gen trên mạch L ( L-strand ) .
Ty thể có chứa hệ gen của riêng chúng, gọi là DNA ty thể (mitochondrial DNA), viết tắt mtDNA; một dấu hiệu cho thấy bào quan này có nguồn gốc từ vi khuẩn thông qua hiện tượng nội cộng sinh (xem Nguồn gốc và con đường tiến hóa ở trên). Tuy nhiên, hệ gen nội cộng sinh tổ tiên của chúng đã mất đi phần lớn các gen, dẫn đến hệ gen ty thể hiện nay là một trong những hệ gen thoái hóa mạnh nhất khi tính trên cấp độ sinh vật.
Hệ gen ty thể người là một phân tử DNA vòng mang khoảng 16 kilobase.[105] Thông tin di truyền tập hợp trong 37 gen: 13 mã hóa các tiểu đơn vị thuộc những phức hệ hô hấp I, III, IV và V; 22 mã hóa tRNA ty thể (gồm 20 loại tRNA tham gia vận chuyển 20 loại amino acid tiêu chuẩn, cộng với một gen bổ sung mã hóa leucine và serine) và 2 mã hóa rRNA.[105] Mỗi một ty thể có thể chứa từ 2 đến 10 bản sao DNA phát sinh từ phân tử gốc.[106]
Trong khi ở sinh vật nhân sơ, hiện diện một tỷ lệ rất cao DNA mã hóa và vắng đi những đoạn lặp lại. Các gen ty thể được phiên mã theo phương thức phiên mã đa gen (multigenic transcript), sau đó bị cắt nối và bổ sung đuôi poly(A) nhằm hoàn thiện tạo nên mRNA trưởng thành. Không phải tất cả protein cần cho chức năng ty thể đều mã hóa bởi hệ gen ty thể; thực chất phần lớn trong số chúng lại do gen nhân tế bào mã hóa và nhập cảng vào bào quan.[46] Con số chính xác những gen mã hóa thuộc hệ gen nhân và ty thể khác nhau tùy vào từng loài. Hầu hết hệ gen ty thể đều là dạng vòng, ngoại trừ một số ngoại lệ.[107] Chung quy lại, DNA ty thể thiếu sót các intron, như trường hợp hệ gen ty thể con người;[46] tuy một vài intron từng được tìm thấy trong số ít DNA ty thể nhân thực,[108] ví dụ nấm men[109] và sinh vật nguyên sinh,[110] như loài Dictyostelium discoideum.[111] Giữa những vùng mã hóa protein có mặt các phân tử tRNA. Trong suốt quá trình phiên mã, tRNA đạt được hoạt tính cấu dạng L (L-shape) để các enzyme đặc hiệu có thể nhận diện và cắt nối. Gen tRNA ty thể có những đoạn trình tự khác biệt so với tRNA nhân nhưng lại có vẻ giống với tRNA ty thể tìm thấy trong các nhiễm sắc thể nhân với mật độ tương đồng trình tự cao.[112]
Ở động vật, hệ gen ty thể thường là một nhiễm sắc thể vòng đơn đặc trưng, dài xấp xỉ 16 kb và chứa 37 gen. Các gen được bảo tồn tốt có thể sắp xếp đa dạng ở nhiều vị trí. Kỳ lạ thay, mô hình này lại không thể tìm thấy ở loài rận thân thể người (human body louse, Pediculus humanus). Thay vào đó, hệ gen ty thể của nó được phân bổ thành 18 nhiễm sắc thể vòng nhỏ (minicircular chromosome), mỗi một phân tử dài khoảng 3–4 kb và chỉ mang 1 đến 3 gen.[113] Mô hình dẫn xuất này cũng có mặt trong những loài rận hút (sucking louse) khác, nhưng lại không có ở rận nhai (chewing louse). Người ta cũng quan sát thấy quá trình tái tổ hợp diễn ra giữa các nhiễm sắc thể nhỏ. Nguyên nhân chính xác của sự khác biệt này vẫn chưa thể hiểu rõ.
Hệ thống mã di truyền thay thế sửa chữa[sửa|sửa mã nguồn]
Ngoài những biến thể nhẹ của bộ mã di truyền tiêu chuẩn đã Dự kiến từ trước, [ 114 ] người ta không còn phát hiện thêm bất kể mã nào cho đến năm 1979, khi những nghiên cứu và điều tra trên những gen ty thể người cho rằng chúng có sử dụng một bộ mã sửa chữa thay thế. [ 115 ] Tuy vậy, ty thể của nhiều sinh vật nhân thực khác, gồm có hầu hết thực vật, vẫn sử dụng mạng lưới hệ thống mã di truyền tiêu chuẩn. [ 116 ] Từ đó trở đi, nhiều biến thể nhỏ của bộ mã liên tục được phát hiện, [ 117 ] gồm cả những mã thay thế sửa chữa trong ty thể. [ 118 ] Thêm nữa, ba codon AUA, AUC và AUU đều là những codon khởi đầu .
| Sinh vật | Codon | Hệ thống tiêu chuẩn | Hệ thống ty thể |
|---|---|---|---|
| Động vật có vú | AGA, AGG | Arginine | Codon kết thúc |
| Động vật không xương sống | AGA, AGG | Arginine | Serine |
| Nấm | CUA | Leucine | Threonine |
| Chung ba nhóm trên | AUA | Isoleucine | Methionine |
| UGA | Codon kết thúc | Tryptophan |
Một vài sai khác trên được coi là giả đổi khác trong bộ mã di truyền do hiện tượng kỳ lạ chỉnh sửa RNA, xảy ra phổ cập trong ty thể. Ở thực vật bậc cao, người ta nghĩ rằng CGG mã hóa cho tryptophan, không phải arginine ; tuy nhiên, codon phát hiện trên RNA sau tinh sửa lại là UGG, tương thích với mạng lưới hệ thống mã di truyền tiêu chuẩn trong mã hóa tryptophan. [ 119 ] Đáng chú ý quan tâm là hệ mã di truyền ty thể động vật hoang dã chân đốt đã trải qua sự kiện tiến hóa song hành nội ngành, dẫn đến vẫn sống sót một vài sinh vật duy nhất dịch mã AGG ra lysine. [ 120 ]
Tiến hóa và sự phong phú[sửa|sửa mã nguồn]
Hệ gen ty thể chứa lượng gen ít hơn rất nhiều so với vi khuẩn, tổ tiên của chúng. Bên cạnh một số gen đã hoàn toàn mất đi, có thể vẫn còn ít gen được chuyển đến nhân, như gen mã hóa những tiểu đơn vị protein thuộc Phức hệ II trong hô hấp tế bào.[105] Người ta cho rằng sự kiện này tương đối phổ biến xuyên suốt thời gian tiến hóa. Một vài sinh vật, như Cryptosporidium, có loại ty thể thiếu sót DNA, phỏng chừng tất cả các gen của ty thể đó khả năng đã mất hoặc bị chuyển đi.[121] Trong Cryptosporidium, ty thể có hệ thống sản xuất ATP được tu sửa, cho thấy ký sinh trùng này đã tiến hóa chống lại nhiều tác nhân ức chế ty thể thường gặp như cyanide, azua và atovaquone.[121]
Nhân đôi và di truyền[sửa|sửa mã nguồn]
Ty thể phân cắt bằng cách phân đôi, tựa như phân bào ở vi trùng. [ 122 ] Sự điều hòa hoạt động giải trí phân đôi ty thể độc lạ giữa những sinh vật nhân thực. Trong nhiều tổ chức triển khai nhân thực đơn bào, sự sinh trưởng và phân loại bào quan này phối hợp với chu kỳ luân hồi tế bào. Tỷ dụ, một ty thể đơn có năng lực phân loại đồng điệu song song với nhân. Quá trình phân đôi và phân phối này phải được trấn áp ngặt nghèo để mỗi tế bào con đều nhận được tối thiểu một ty thể. Trong những sinh vật nhân thực khác ( như thú ), hoàn toàn có thể phần nhiều ty thể liên tục sao chép DNA của chúng và phân đôi nhằm mục đích cung ứng nhu yếu nguồn năng lượng tế bào, hơn là phải hoạt động giải trí đồng pha với chu kỳ luân hồi tế bào. Một khi nhu yếu nguồn năng lượng tăng cao, ty thể sẽ tích cực sinh trưởng và phân đôi. Còn khi nhu yếu nguồn năng lượng xuống thấp, ty thể bị phân hủy hay trở nên bất hoạt. Theo những ví dụ trên, và tương phản với thực trạng ở nhiều sinh vật nhân thực đơn bào, rõ ràng ty thể được phân phối ngẫu nhiên cho những tế bào con trong quá trình phân loại tế bào chất. Hiểu biết về hoạt độ động lực ty thể, tựa như tính cân đối giữa sự dung hợp và phân đôi, đã cho thấy những đổi khác công dụng và cấu trúc trong hình thái học ty thể là tác nhân trọng điểm của bệnh lý học, tương quan đến một số ít cơ sở khởi phát bệnh. [ 123 ]
Những lý thuyết về quá trình phân đôi ty thể đạt được độ tin cậy bằng việc trực quan hóa nhờ kính hiển vi huỳnh quang và kính hiển vi điện tử truyền qua tiêu chuẩn (transmission electron microscopy, TEM). Ta phải sử dụng tận hai kính là do độ phân giải kính hiển vi huỳnh quang (~200 nm) không đủ để phân biệt chi tiết các cấu trúc, như màng kép ty thể trong phân đôi bào quan hay thậm chí nhận ra từng ty thể riêng biệt trong trường hợp nằm gần nhau. Mặc dù TEM tiêu chuẩn vẫn còn một số hạn chế kỹ thuật[cái gì?] trong việc xác minh sự phân đôi ty thể. Gần đây người ta cũng sử dụng phương pháp chụp cắt lớp điện tử lạnh (cryo-electron tomography) để trực quan hóa phân đôi ty thể trên mẫu tế bào ngậm nước đông lạnh nguyên vẹn. Phương pháp này tiết lộ rằng ty thể phân đôi bằng cách nảy chồi.[124]
Cơ chế di truyền của những gen ty thể trong từng đơn vị chức năng bào quan riêng không liên quan gì đến nhau không như những gen nhân tế bào. Thông thường, mỗi ty thể chỉ thừa kế thông tin di truyền từ một bên cha mẹ đơn nhất. Ở người, khi một tế bào trứng được tinh trùng thụ tinh, nhân tế bào trứng và tinh trùng sẽ cùng góp chung một lượng vật chất di truyền như nhau để tạo nên nhân tế bào hợp tử. Trong khi ở ty thể lại ngược lại, DNA ty thể thường chỉ đến từ tế bào trứng. Ty thể tinh trùng cũng nhập bào vào trứng, nhưng không góp được thông tin di truyền nào cho phôi. [ 125 ] Nguyên nhân là do những ty thể từ bố bị ubiquitin ghi lại và phôi sẽ cho tiêu hủy sau này. [ 126 ] Tế bào trứng chứa tương đối ít ty thể, nhưng nhóm ty thể này lại sống sót và liên tục phân đôi để cư trú trong những tổ chức triển khai sống trưởng thành. Qua đó, hầu hết trường hợp giao phối chỉ di truyền ty thể từ mẹ, khát quát lên thành khái niệm di truyền dòng mẹ. Phương thức di truyền này xuất hiện trong hầu hết sinh vật, gồm có hầu hết động vật hoang dã. Nhưng nhiều lúc cũng có trường hợp di truyền ty thể từ bố ; Open nhất định ở một số ít loài cây ngành Thông, dù không gồm có chi Thông và chi Taxus. [ 127 ] Còn ở họ Vẹm, di truyền dòng bố chỉ xảy ra trong con đực. [ 128 ] [ 129 ] [ 130 ] Người ta cũng cho rằng tỷ suất Open phương pháp di truyền này ở con người là rất thấp. [ 131 ] Một quan điểm gần đây trình diễn việc ty thể sẽ gây rút ngắn tuổi thọ giống đực, nguyên do là do chúng chỉ được thừa kế từ mẹ. Ngược lại, tiến trình tinh lọc tự nhiên ở cỏ dại lại cho thấy ty thể giảm mất thời cơ sống sót của giống cái, và như vậy ty thể ít có năng lực truyền đến những thế hệ tiếp theo. Kết luận, từ những dẫn chứng trên đã cho tất cả chúng ta thấy một trong những nguyên do dẫn đến hiện tượng kỳ lạ giống cái loài người và động vật hoang dã có khuynh hướng sống lâu hơn giống đực. Các tác giả của đánh giá và nhận định trên chứng minh và khẳng định đây là một lý giải chưa tổng lực. [ 132 ]Di truyền một bên cha mẹ tạo ra ít thời cơ tái tổng hợp di truyền giữa những dòng ty thể khác nhau, mặc dù rằng mỗi ty thể đơn hoàn toàn có thể chứa từ 2 – 10 bản sao DNA. [ 106 ] Vì nguyên do này, người ta thường nghĩ DNA ty thể sản sinh theo phương pháp phân cắt thành hai ( phân đôi ). Có thể do sự tái tổng hợp ưu tiên bảo tồn toàn vẹn hệ di truyền hơn là duy trì tính phong phú. Dù vậy, đã có những điều tra và nghiên cứu bật mý dẫn chứng quy trình tái tổng hợp có xảy ra ở DNA ty thể. Theo như trên, rõ ràng những enzyme triển khai tái tổng hợp có hiện hữu trong tế bào động vật hoang dã có vú. [ 133 ] Hơn nữa, những chứng cứ còn gợi ý ty thể động vật hoang dã hoàn toàn có thể là hệ quả của quy trình tái tổng hợp từ trước. [ 134 ] Dữ liệu tương tự như ở người thì còn gây tranh cãi, dù những dẫn chứng gián tiếp vẫn cho thấy có sống sót sự tái tổng hợp. [ 135 ] [ 136 ] Nếu tái tổng hợp không diễn ra, hàng loạt trình tự DNA ty thể sẽ trở thành một haplotype duy nhất, khiến trường hợp này có ích trong việc điều tra và nghiên cứu lịch sử dân tộc tiến hóa quần thể .Những ty thể được di truyền theo một bên cha mẹ và ít hoặc không tái tổng hợp năng lực sẽ chịu tác động ảnh hưởng từ hiệu ứng chốt hãm ngược Muller ( Muller’s ratchet ), tích tụ vô tình những đột biến có hại cho đến khi công dụng bào quan trọn vẹn biến mất. Quần thể ty thể động vật hoang dã tránh đi sự tích tụ này trải qua một quy trình tiến hóa gọi là nút cổ chai ty thể ( mitochondrial bottleneck ). Nút cổ chai tận dụng những quy trình ngẫu nhiên hiện hữu trong tế bào để ngày càng tăng sự chênh lệch hàm lượng nạp tải đột biến giữa những tế bào theo tiến trình tăng trưởng sinh vật, như khi : một tế bào trứng đơn chứa một lượng mtDNA đột biến hoàn toàn có thể trở thành một phôi mang nhiều tế bào khác nhau có lượng nạp tải đột biến độc lạ. Sau đó hoàn toàn có thể diễn ra sự tinh lọc Lever tế bào nhằm mục đích vô hiệu những tế bào chứa nhiều mtDNA đột biến, dẫn đến không thay đổi hóa hay giảm thiểu hàm lượng nạp tải đột biến giữa những thế hệ. Cơ chế đằng sau quy trình nút cổ chai vẫn còn đang tranh luận. [ 137 ] [ 138 ] [ 139 ] Một nghiên cứu và điều tra tổng hợp toán học lẫn thực nghiệm gần đây đã phân phối vật chứng về sự tổng hợp trong phân vùng ngẫu nhiên mtDNA khi phân bào và thay đổi ngẫu nhiên những phân tử mtDNA trong tế bào. [ 140 ]
Nghiên cứu di truyền quần thể[sửa|sửa mã nguồn]
Việc DNA ty thể gần như không xảy ra sự tái tổng hợp di truyền khiến nó trở thành một nguồn thông tin có ích cho những nhà khoa học nghiên cứu và điều tra về di truyền học quần thể và sinh học tiến hóa. [ 141 ] Vì tổng thể DNA ty thể đều di truyền ở dạng phân tử đơn, hay haplotype, nên tất cả chúng ta hoàn toàn có thể trực quan hóa mối quan hệ DNA ty thể giữa những thành viên khác nhau thành một cây gen ( cây phát sinh chủng loại ). Những hình mẫu của cây gen hoàn toàn có thể được sử dụng để suy luận lịch sử vẻ vang tiến hóa quần thể. Ví dụ nổi bật là khi ta vận dụng điều này trong di truyền học tiến hóa loài người, bằng cách sử dụng giải pháp đồng hồ đeo tay phân tử để xác lập thời hạn sống sót tổ tiên Eve ty thể gần nhất. [ 142 ] [ 143 ] Người ta thường cho rằng suy luận này ủng hộ can đảm và mạnh mẽ so với quy trình lan rộng ra chủ quyền lãnh thổ khỏi châu Phi gần đây của loài người văn minh. [ 144 ] Một ví dụ khác cũng về con người là việc giải trình tự DNA ty thể từ xương người Neanderthal. Qua đó thấy được, khoảng cách tiến hóa tương đối lớn giữa những trình tự DNA ty thể của người Neanderthal và người tân tiến là vật chứng cho sự thiếu đi hoạt động giải trí giao phối giữa hai loài người trên. [ 145 ]Tuy vậy, DNA ty thể chỉ phản ánh lịch sử dân tộc tiến hóa của giống cái trong quần thể và do đó không hề đại diện thay mặt cho lịch sử vẻ vang tiến hóa quần thể chung của hàng loạt thành viên. Việc này hoàn toàn có thể khắc phục bằng cách sử dụng những trình tự di truyền từ bố, ví dụ như vùng không tái tổng hợp trên nhiễm sắc thể Y. [ 144 ] Còn ở một ngữ nghĩa tổng quát hơn, chỉ những điều tra và nghiên cứu mà bao hàm cả DNA nhân mới hoàn toàn có thể cung ứng cho tất cả chúng ta một lịch sử vẻ vang tiến hóa quần thể tổng lực. [ 146 ]Những đo đạc gần đây từ kỹ thuật đồng hồ đeo tay phân tử trên DNA ty thể [ 147 ] đã báo cáo giải trình cứ mỗi 7884 năm lại Open 1 đột biến, kể từ niên đại sống sót tổ tiên chung gần nhất của loài người và vượn, điều này thích hợp với tần số đột biến DNA nhiễm sắc thể thường ( 10 − 8 trên một base trên một thế hệ [ 148 ] ) .
Rối loạn và bệnh lý[sửa|sửa mã nguồn]
Bệnh lý ty thể[sửa|sửa mã nguồn]
Sự rối loạn tổn thương và tiến triển trong bào quan ty thể là tác nhân quan trọng gây ra hàng loạt căn bệnh ở người, nguyên do là do những tác động ảnh hưởng của chúng đến quy trình trao đổi chất tế bào. Rối loạn ty thể thường biểu lộ triệu chứng như một dạng rối loạn thần kinh, gồm có tự kỷ. [ 13 ] Rối loạn cũng hoàn toàn có thể biểu lộ thành bệnh cơ, tiểu đường, nhiều bệnh nội tiết và một loạt những bệnh lý mạng lưới hệ thống khác. [ 149 ] Đột biến mtDNA thì gây ra hội chứng Kearns – Sayre, hội chứng MELAS và bệnh teo thị thần kinh Leber. [ 150 ] Trong hầu hết trường hợp, những bệnh này được di truyền từ mẹ sang con, khi mà hợp tử thu nhận ty thể và dẫn nhập mtDNA từ tế bào trứng. Những bệnh lý như hội chứng Kearns – Sayre, hội chứng Pearson và liệt mắt ngoại lai tuần tiến mạn tính được cho là khởi phát từ quy trình tái sắp xếp mtDNA quy mô lớn, trong khi những bệnh khác như hội chứng MELAS, teo thị thần kinh Leber, hội chứng động kinh giật cơ với sợi cơ không đều ( myoclonic epilepsy with ragged red fibers, MERRF ), v.v. lại xuất phát từ những đột biến điểm trên mtDNA. [ 149 ]Ở 1 số ít bệnh, những khiếm khuyết trên gen nhân là tác nhân dẫn đến rối loạn tính năng protein ty thể. Bao gồm những bệnh : mất điều hòa Friedreich ( hay thất điều Friedreich ), liệt hai chi dưới co cứng di truyền ( hereditary spastic paraplegia ) và bệnh Wilson. [ 151 ] Những bệnh trên đều di truyền theo quan hệ tính trạng trội lặn, tương tự như như hầu hết bệnh lý di truyền khác. Một loạt những rối loạn hoàn toàn có thể gây ra từ những loại sản phẩm enzyme phosphoryl hóa oxy hóa sản sinh từ những đột biến tại nhân, ví dụ bệnh thiếu vắng coenzyme Q10 và hội chứng Barth. [ 149 ] Những tác động ảnh hưởng từ môi trường tự nhiên cũng có năng lực tương tác với những tố bẩm di truyền ( hereditary predisposition ) và khởi phát bệnh lý ty thể. Ví dụ, hoàn toàn có thể có một mắc xích giữa việc phơi nhiễm thuốc trừ sâu và phát bệnh Parkinson sau đó. [ 152 ] [ 153 ] Những bệnh lý khác có căn nguyên từ sự rối loạn ty thể còn gồm có : tinh thần phân liệt, rối loạn lưỡng cực, suy giảm trí nhớ, bệnh Alzheimer, [ 154 ] bệnh Parkinson, động kinh, đột quỵ, bệnh tim mạch, hội chứng căng thẳng mệt mỏi mãn tính, viêm võng mạc sắc tố và đái tháo đường. [ 155 ] [ 156 ]Căng thẳng oxy hóa trung gian qua ty thể là một tác nhân gây ra bệnh cơ tim ở bệnh nhân tiểu đường típ 2. Sự ngày càng tăng dẫn truyền axit béo đến tim khiến cơ quan này phải tăng cường hấp thu những axit trải qua những tế bào cơ tim, dẫn đến hệ quả tăng cường quy trình oxy hóa axit béo trong nhóm tế bào này. Tiến trình oxy hóa sẽ làm tăng hàm lượng đương lượng khử sẵn có để chu cấp cho chuỗi chuyền điện tử ở ty thể, và ở đầu cuối gây nên sự tăng sinh nồng độ những chủng chất hoạt động giải trí có oxy ( reactive oxygen species, ROS ). Tiếp theo, ROS sẽ khiến tăng lượng protein tách cặp ( uncoupling protein, UCP ) và tăng nhanh rò rỉ proton qua những chất mang adenine nucleotide translocator ( ANT ). Các hoạt động giải trí tổng hợp trên khiến thoái biến thế năng màng ty thể và được gọi là hiện tượng kỳ lạ tách cặp. Tiếp sau, sự tách cặp còn khiến ty thể tăng hiệu suất tiêu thụ oxy, làm tồi tệ thêm quy trình oxy hóa axit béo. Tạo ra một vòng tách cặp luẩn quẩn ; hơn nữa, dù cho hiệu suất tiêu thụ oxy tăng cao, nhưng hoạt động giải trí sản xuất ATP lại không tăng lên tương ứng do ty thể đã bị tách cặp. Vì thế, lượng ATP khả dụng tiêu nhỏ là hậu quả tận cùng trong tiến trình thâm hụt nguồn năng lượng, bộc lộ qua sự suy biến tính năng tim và rối loạn năng lực co bóp. Nghiêm trọng hơn, những tiến trình trên còn khiến lưới cơ tương hư hỏng tính năng giải phóng calci và làm ty thể hạn chế năng lực đệm hấp những cao điểm nồng độ calci trong bào tương, một yếu tố quan trọng để truyền tải thông suốt tín hiệu ion khi co cơ. Sự sụt giảm nồng độ calci nội ty thể sẽ làm tăng kích hoạt những enzyme dehydrogenase và sinh tổng hợp ATP. Vì vậy, bên cạnh việc giảm sút do sự oxy hóa axit béo, hoạt động giải trí tổng hợp ATP còn bị suy yếu do nghèo hóa tín hiệu calci, gây ra những biến chứng tim mạch so với bệnh nhân tiểu đường. [ 157 ]
Những mối liên hệ khả thi đến tiến trình lão hóa[sửa|sửa mã nguồn]
Với công dụng là xí nghiệp sản xuất nguồn năng lượng của tế bào, nơi liên tục xảy ra những phản ứng oxy hóa, đôi lúc ty thể cũng bị rò rỉ những electron cao nguồn năng lượng trong chuỗi chuyền điện tử rồi dẫn đến hình thành những chủng chất hoạt động giải trí có oxy ( reactive oxygen species, ROS ). Đây được xem là hệ quả của thực trạng căng thẳng mệt mỏi oxy hóa đáng kể trong ty thể chứa bộ mtDNA có tỷ suất đột biến cao. [ 158 ] Mối liên hệ mắc xích giữa sự lão hóa và căng thẳng mệt mỏi oxy hóa không hẳn là mới và đã được đề xuất kiến nghị năm 1956, [ 159 ] sau đó khái quát thành triết lý gốc tự do ty thể của sự lão hóa. [ 160 ] Một vòng luẩn quẩn Open, đó là khi căng thẳng mệt mỏi oxy hóa khởi phát những đột biến trên DNA ty thể, từ đó sản sinh những enzyme không bình thường và lại liên tục gây ra căng thẳng mệt mỏi oxy hóa .Một số biến hóa trên ty thể hoàn toàn có thể diễn ra trong suốt tiến trình lão hóa. [ 161 ] Phân tích mẫu mô từ những bệnh nhân cao tuổi cho thấy sự sụt giảm hoạt tính enzyme của những protein thuộc chuỗi chuyền điện tử. [ 162 ] Tuy vậy, mtDNA đột biến chỉ xuất hiện trong khoảng chừng 0,2 % tế bào thật sự già. [ 163 ] Các nhà khoa học đưa ra triết lý cho rằng hiện tượng kỳ lạ khuyết đoạn quy mô lớn hệ gen ty thể đã dẫn đến căng thẳng mệt mỏi oxy hóa mức độ cao và phá chết neuron ở bệnh nhân Parkinson. [ 164 ]
Ty thể trong văn hóa truyền thống đại chúng[sửa|sửa mã nguồn]

 khoa học viễn tưởng A Wind in the Door và Parasite Eve có những miêu tả hư cấu về bào quan ty thể, là thành tố chính của cốt truyện.Hai cuốn tiểu thuyết vàcó những miêu tả hư cấu về bào quan ty thể, là thành tố chính của diễn biến .
khoa học viễn tưởng A Wind in the Door và Parasite Eve có những miêu tả hư cấu về bào quan ty thể, là thành tố chính của cốt truyện.Hai cuốn tiểu thuyết vàcó những miêu tả hư cấu về bào quan ty thể, là thành tố chính của diễn biến .
Tiểu thuyết khoa học viễn tưởng A Wind in the Door xuất bản năm 1973 của tác giả Madeleine L’Engle có nội dung về bào quan ty thể trong cơ thể nhân vật chính Charles Wallace Murry, được hư cấu hóa thành nơi sinh sống của những sinh vật gọi là farandolae (lấy từ tên một điệu vũ nổi tiếng ở xứ Provence, Pháp). Ngoài ra, cốt truyện cũng xuất hiện những nhân vật đi du lịch bên trong ty thể của Murry.
Tiểu thuyết kinh dị viễn tưởng Parasite Eve xuất bản năm 1995 của tác giả Hideaki Sena mô tả ty thể có thể ý thức tương đối và có khả năng điều khiển tâm trí, nỗ lực lợi dụng những đặc tính đó để vượt trên sinh vật nhân thực, trở thành dạng sống ưu thế. Tiểu thuyết này đã được chuyển thể thành bộ phim, video game và video game nối tiếp cùng tên, ăn nhập với cốt truyện ban đầu.
Trong vũ trụ giả tưởng Chiến tranh giữa các vì sao, những vi sinh vật ám chỉ với tên “midi-chlorian” có khả năng cảm giác và sử dụng Thần lực. Trong bộ phim năm 1999 Chiến tranh giữa các vì sao (Phần I): Bóng ma đe dọa, giới thiệu lần đầu những sinh vật midi-chlorian, đạo diễn George Lucas đã miêu tả chúng như “một phác họa mơ hồ từ ty thể”.[165] Sau này, các nhà phân loại học lấy ý tưởng từ tên của nhóm sinh vật hư cấu trên để đặt ra tên chi Midichloria thuộc giới Vi khuẩn.
Trong môi trường tự nhiên giáo dục những môn khoa học, ty thể thường thẩm thấu đều đặn vào trí óc học viên với cụm từ nổi tiếng ” ty thể là xí nghiệp sản xuất nguồn năng lượng của tế bào “, tạo ra hệ quả là bộc phát thành một hình tượng meme phổ cập trên Internet. [ 166 ] Meme này ý niệm rằng những giá trị trọng tâm của nền giáo dục trung học đã lệch khỏi nhu yếu đảm nhiệm kiến thức và kỹ năng sống, cho rằng lượng kiến thức và kỹ năng học thuật này, như hiểu biết về vai trò của ty thể, tương đối không có quyền lợi trong thực tiễn. [ 167 ]
Tuyên bố chung
Xem thêm: Bột Yến Mạch Nguyên Chất Giá Bao Nhiêu Tiền 1Kg? Yến Mạch Úc Tươi Nguyên Chất Nguyên Cán Loại 1
 Bài viết này có sử dụng các nguyên liệu từ Science Primer phát hành tại NCBI, vì đây là một tác phẩm của chính quyền liên bang Hoa Kỳ nên nó thuộc phạm vi công cộng [3].
Bài viết này có sử dụng các nguyên liệu từ Science Primer phát hành tại NCBI, vì đây là một tác phẩm của chính quyền liên bang Hoa Kỳ nên nó thuộc phạm vi công cộng [3].
Liên kết ngoài[sửa|sửa mã nguồn]
Source: https://trangdahieuqua.com
Category: Làm đẹp


